
 Автор:
Автор: Н.М. Чернова, А.М. Былова
Общая экология. Учебник
М.: Дрофа, 2004
| Предыдущая |
Содержание статьи:
Глава 3. Важнейшие абиотические факторы и адаптации к ним организмов
3.3. Влажность
3.3.3. Водный баланс наземных животных
Животные получают воду тремя основными путями: через питье, вместе с сочной пищей и в результате метаболизма, т. е. за счет окисления и расщепления органических веществ – жиров, белков и углеводов.
Некоторые животные могут впитывать воду через покровы из влажного субстрата или воздуха, например личинки некоторых насекомых – мучного хрущака, жуков-щелкунов и др.
Потери воды у животных происходят через испарение покровами или со слизистых оболочек дыхательных путей, путем выведения из тела мочи и непереваренных остатков пищи.
Хотя животные могут выдерживать кратковременные потери воды, но в целом расход ее должен возмещаться приходом. Потери воды приводят к гибели скорее, чем голодание.
Виды, получающие воду в основном через питье, сильно зависят от наличия водопоев. Это особенно характерно для крупных млекопитающих. В сухих, аридных районах такие животные совершают иногда значительные миграции к водоемам и не могут существовать слишком далеко от них. В африканских саваннах слоны, антилопы, львы, гиены регулярно посещают водопои (рис. 35). Для куланов Бадхызского заповедника водопои определяют летнее размещение табунов, суточный ритм и поведение животных.

Рис. 35. Слоны на водопое
В питьевой воде нуждаются и многие птицы. Ласточки и стрижи пьют на лету, проносясь над поверхностью водоема. Рябки в пустынях ежедневно совершают многокилометровые перелеты к водопоям и приносят воду птенцам. Самцы рябков используют исключительный в своем роде способ переноса воды – они пропитывают ею оперение на груди, а птенцы отжимают клювами набухшие перья.
В то же время многие животные могут обходиться совсем без питьевой воды, получая влагу иными способами.
Влажность воздуха также очень важна для животных, так как от нее зависит величина испарения с поверхности тела (рис. 36). Потери воды через испарение обусловлены также строением покровов. Некоторые виды не могут обитать в сухом воздухе и нуждаются в полном насыщении его водяными парами. Другие без вреда для себя населяют самые засушливые районы.
Среди ряда групп животных можно выделить гигрофилов и ксерофилов, т. е. влаголюбивые и сухолюбивые виды. Промежуточную группу составляют мезофилы. Среди насекомых, например, гигрофильны кровососущие комары, которые активны преимущественно в вечерние и утренние часы, а днем – либо в пасмурную погоду, либо только в тени, под пологом леса, т. е. при повышенной влажности воздуха. Ксерофильны жуки-скакуны, пустынные жуки-чернотелки, пустынная саранча и др.
Способы регуляции водного баланса у животных разнообразнее, чем у растений. Их можно разделить на поведенческие, морфологические и физиологические.
К числу поведенческих приспособлений относятся поиски водопоев, выбор мест обитания, рытье нор и т. п. В норах влажность воздуха приближается к 100 %, даже когда на поверхности очень сухо. Это снижает необходимость испарения через покровы, экономит влагу в организме.
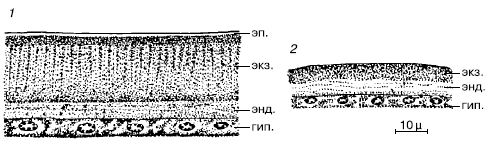
Рис. 36. Строение кутикулы членистоногих (по М. С. Гилярову, 1970):
1– жук мучной хрущах Tenebrio molitor;
2– ветвистоусый рачок Cyclops sp.: гип. – гиподерма, энд. – эндокутикула; экз. – экзокутикула, эп. – эпикутикула
В эффективности поведенческих приспособлений для обеспечения водного баланса можно убедиться на примере пустынных мокриц. Мокрицы – типичные ракообразные, не отличающиеся особыми анатомо-морфологическими приспособлениями к наземному образу жизни. Тем не менее представители рода Hemilepistus освоили самые сухие и жаркие места на Земле – глинистые пустыни. Там они роют глубокие вертикальные норки, где всегда влажно, и покидают их, выходя на поверхность лишь в те часы суток, когда высока влажность приземного слоя воздуха. Когда почва иссушается особенно сильно и возникает угроза снижения влажности воздуха в норке, самки закрывают отверстие сильно склеротизованными передними сегментами тела, создавая замкнутое, насыщенное парами пространство и оберегая молодь от высыхания.
К морфологическим способам поддержания нормального водного баланса относятся образования, способствующие задержанию воды в теле: раковины наземных улиток, ороговевшие покровы рептилий, развитие эпикутикулы у насекомых и т. п. У пустынных жуков-чернотелок срастаются и прирастают к телу надкрылья, вторая пара крыльев редуцируется и между телом и надкрыльями образуется камера, куда выходят дыхальца насекомого. Эта камера открывается наружу лишь небольшой узкой щелью, воздух в ней насыщен водяными парами. Части тела, соприкасающиеся с внешней средой, защищены непроницаемой для воды эпикутикулой.
Физиологические приспособления к регуляции водного обмена – это способность к образованию метаболической влаги, экономии воды при выделении мочи и кала, развитие выносливости к обезвоживанию организма, величина потоотделения и отдачи воды со слизистых.
Выносливость к обезвоживанию, как правило, выше у животных, подвергающихся тепловым перегрузкам. Для человека потеря воды, превышающая 10 % массы тела, смертельна. Верблюды переносят потери воды до 27 %, овцы – до 23, собаки – до 17 %.
Экономия воды в пищеварительном тракте достигается всасыванием воды кишечником и продуцированием сухого кала. Содержание воды в испражнениях животных варьирует в зависимости от состава корма, но в целом отражает приспособленность к обитанию в разных условиях влажности. Например, на 100 г сухого помета коров на пастбище приходится 566 г воды, тогда как у верблюдов – 109, а при безводной диете – всего 76 г.
У насекомых, обитающих в аридных районах, выделительные органы – мальпигиевы сосуды – свободными концами входят в тесный контакт со стенкой задней кишки и всасывают воду из ее содержимого. Таким образом вода вновь возвращается в организм (пустынные жуки-чернотелки, муравьиные львы, личинки божьих коровок и др.).
Для экономии воды, выводимой через почки, нужна перестройка азотного обмена. При распаде белков у большинства водных организмов образуется аммиак, который токсичен для цитоплазмы даже в малых концентрациях. На процесс его образования и выведения тратится много воды. У наземных животных аммиак присутствует среди продуктов обмена только у тех форм, которые обитают в условиях достаточного обеспечения водой, например у тлей, непрерывно питающихся соком растений. Основной компонент выделяемой мочи у наземных млекопитающих – мочевина. Это менее токсичный продукт обмена, который может накапливаться в плазме и полостных жидкостях и выводиться в более концентрированных растворах, что экономит воду. С мочой выводятся также различные соли. Общая концентрация мочи по сравнению с плазмой может служить показателем способности к экономии воды при экскреции. У человека моча концентрированнее плазмы в 4,2 раза, у овец – в 7,6, у верблюда – в 8, у тушканчиков – в 14 раз.
Чешуйчатые пресмыкающиеся и сухопутные черепахи – группы, освоившие наиболее аридные районы, – выделяют малорастворимую мочевую кислоту. Это же характерно для птиц и высших насекомых. Паукообразные выделяют гуанин. При образовании гуанина и мочевой кислоты затрачивается минимальное количество воды.
Жизнь за счет метаболической влаги доступна не всем животным. Окисление жиров требует большого количества кислорода, а дополнительная вентиляция легких в сухом воздухе сопровождается потерей водяных паров. Жир в горбах верблюдов не является для них основным источником водоснабжения, так как расход воды на усиленное дыхание при терморегуляции равен или даже превышает количество получаемой метаболической воды. Поэтому верблюды нуждаются в периодическом питье.
Мелкие млекопитающие, спасающиеся от жары в прохладных норах, могут покрывать значительную часть своих расходов воды в результате окислительных процессов, так как им не требуется дополнительно вода на терморегуляцию. Почти исключительно на сухом корме живут такие пустынные виды, как многие тушканчики, американская кенгуровая крыса, африканская песчанка и др. (рис. 37). Кенгуровых крыс содержали в лаборатории на сухой перловой крупе. При этом из 100 г корма, потребляемого зверьком за месяц, образуется около 54 г воды. Кроме нее, животные использовали лишь абсорбированную крупой влагу, содержание которой, в зависимости от влажности воздуха, составляет от 10 до 18 %.

Рис. 37. Животные, способные жить за счет метаболической влаги: 1 – пустынный мешотчатый прыгун; 2 – карликовая песчанка; 3 – жирнохвостый тушканчик (по В. Е. Соколову и др., 1977)
Метаболическую воду в большей мере, чем позвоночные животные, могут использовать насекомые, так как трахейная система насекомых осуществляет эффективный воздушный дренаж с малыми потерями на испарение. У многих видов жировое тело служит преимущественно источником воды, а не энергетических запасов. Гусеницы платяной моли, мельничной огневки, амбарный и рисовый долгоносики и многие другие живут исключительно за счет сухой пищи.
Испарение, связанное с необходимостью терморегуляции, может служить причиной истощения водных ресурсов организма. В пустынях противостоять перегреву путем испарения воды могут только крупные животные. Общая тепловая нагрузка пропорциональна относительной поверхности и поэтому особенно велика для мелких форм. Для животного массой 100 г расход воды составил бы в час около 15 % от массы тела, а массой 10 г – около 30 %, т. е. за немногие часы была бы истрачена вся вода организма. Поэтому мелкие гомойотермные животные в сухом и жарком климате избегают воздействия жары и экономят влагу, укрываясь под землей.
У пойкилотермных повышение температуры тела вслед за нагреванием воздуха позволяет избегать излишних потерь воды, которая тратится у гомойотермных для поддержания постоянной температуры.
Преимущества колеблющейся температуры тела используют и животные с хорошей температурной регуляцией, специализированные к жизни в пустыне. Например, верблюды способны отключать на некоторое время терморегуляционное испарение. При этом животное массой 500 кг аккумулирует около 10 500 кДж, для рассеивания которых потребовалось бы затратить 5 л воды. Накопленное тепло выводится из организма ночью путем прямого излучения, когда воздух становится прохладнее тела.
Пойкилотермные животные, однако, не могут полностью избежать потерь воды на испарение. Даже у рептилий с их ороговевшим эпидермисом потери воды через кожу значительны. У мелких ящериц они могут достигать 20 % и более от массы тела за сутки. Поэтому и для пойкилотермных основной путь сохранения водного баланса при жизни в пустыне – это избегание излишних тепловых нагрузок.
| Предыдущая |