
 Автор:
Автор: И.С. Белюченко, О.А. Мельник
Сельскохозяйственная экология
Учебное пособие. – Краснодар: Изд-во КГАУ, 2010. — 297 с.
| Предыдущая |
Содержание статьи:
2. Теоретические и практические аспекты развития сельскохозяйственной экологии
2.4. Взаимоотношения в агросистемах
Агросистема представляет собою понятие широкое и не имеющее пространственного и временного ранга. Например, совокупность земель большого хозяйства, бригадный стан, животноводческая ферма и т.д. могут рассматриваться как отдельные агросистемы с массой особенностей, свойственных каждому из выделенных формирований. Основу большинства агросистем составляет агрофитоценоз, представляющий собою относительно однородный участок поля с произрастающей на нем растительностью. Задача специалистов сельского хозяйства и экологов – создать агроландшафт с оптимальными свойствами, обусловливающими получение высокого дохода и сохранения природы. Во всех агроландшафтах на вершине пирамиды находится человек, стремящийся получить больше продуктов растениеводства и животноводства.
Оценку эффективности агроландшафтов можно проводить по следующим признакам:
1. Видовое многообразие у однолетних посевов минимально и в основном поддерживается за счет сорных растений и их консортов.
2. Стабильность (относительное постоянство от года к году) поддерживается за счет дополнительных энергетических затрат (поливы, удобрения и т.д.).
3. Гибкость (восстанавливаемость) у однолетних посевов отсутствует для культурного растения и весьма выражена для сорняков.
4. Направленность годичных изменений к устойчивому состоянию; в однолетних посевах этому процессу мешает человек, прерывающий поступательное развитие стадий восстановительной сукцессии.
5. Автономность (способность к самосохранению) при прекращении деятельности человека не свойственна для однолетников и четко проявляется у многолетних сообществ.
Взаимоотношения между организмами в агросистемах весьма разнообразны, но изучены, к сожалению, еще недостаточно. Выделяют следующие основные формы взаимоотношений: симбиотические и антагонистические.
Агроландшафтные системы, независимо от объема, составлены многочисленными видами организмов – от простейших до высших растений и наиболее развитых животных. Организмы в системе не существуют отдельно (сами по себе), а находятся в определенных взаимоотношениях друг с другом (табл.1). Эти взаимоотношения могут быть простыми или сложными, кратковременными или постоянными, прямыми или косвенными.
Таблица 1. Типы взаимоотношений между организмами в экосистеме
|
Форма |
Взаимоотношения |
Виды |
|
|
А |
В |
||
|
Симбиоз |
Мутуализм Комменсализм Промкооперация Нейтрализм |
+ + + 0 |
+ 0 + 0 |
|
Антагонизм |
Антибиоз Аменсализм Паразитизм Хищничество Конкуренция |
0 + + + – |
– – – – – |
Симбиоз. Этим термином определяют такие взаимоотношения между разными организмами, которые, как правило, способствуют их процветанию или, по крайней мере, не наносят никакого вреда ни одному из них. Симбиотические взаимоотношения явились продуктом, очевидно, длительной коэволюции между организмами. Если присутствие одного вида необходимо для существования другого, то их взаимоотношения называют мутуализмом. Классическим примером мутуалистических отношений может быть взаимополезная связь термитов и простейших. Простейшие (жгутиковые) обитают в кишечнике термитов. Последние питаются древесиной, несмотря на то, что они не имеют разрушающего клетчатку фермента. Этот фермент имеют жгутиковые, обитающие в кишечнике термитов. Фермент вызывает распад клетчатки древесины до сахаров. Образовавшиеся сахара частично используются простейшими в процессе обмена веществ, а остальную часть расходуют термиты. Термиты не могут существовать без фауны в кишечнике. Свежевылупившийся термит интенсивно лижет анальное отверстие другого термита, чтобы заполучить жгутиковые. Каждая линька термитов сопровождается потерей жгутиковых. Однако, поскольку термиты живут колониями, каждая особь получает простейших от соседей. Жгутиковым такое соседство тоже выгодно, поскольку оно обеспечивает им пищу всегда в избытке, и кроме того, благоприятные условия местонахождения. Эволюционно такие виды жгутиковых могут жить только в кишечнике живых организмов.
Не менее интересным примером мутуализма являются взаимоотношения между целым рядом видов микроорганизмов – разрушителей клетчатки – и крупным рогатым скотом. Целлюлозобактеры обитают в кишечнике животных, которые потребляют довольно большие количества грубого корма, богатого клетчаткой. Животные практически не могут самостоятельно (без микроорганизмов) перерабатывать клетчатку, поскольку не имеют соответствующих ферментов. Такие ферменты образуют целлюлозобактеры, обитающие в кишечнике животных. Ферменты обусловливают распад клетчатки сена, силоса, зеленой массы и других кормов до сахаров, вырабатывают органические кислоты, переводят неорганический азот в органический и производят различные витамины. Полученные таким образом сахара используются в основном бактериями в качестве энергетического материала, небольшая их часть расходуется организмом животных для формирования новых структур и поддержания жизнедеятельности. В основном животные используют органические кислоты, которые для них являются энергетическим материалом. Кстати, они оказывают также большое влияние на качество молока. Животные не могут существовать без фауны в кишечнике. Маленькие телята, чтобы получить микрофлору, очень рано начинают лизать листочки травы, где некоторые виды микроорганизмов могут встречаться в течение короткого времени, и тесно контактируют с матерью. Микроорганизмам такое соседство весьма выгодно: им оно всегда дает пищу, а также обеспечивает безопасные и наиболее оптимальные условия существования. Такое сосуществование столь далеких друг от друга по уровню развития организмов, безусловно, явилось результатом их сопряженной эволюции.
Мутуалистические взаимоотношения сложились также между бобовыми (высшие растения) и азотфиксирующими бактериями. Так, многие виды бактерий этого рода не способны фиксировать азот воздуха сами по себе, но великолепно осуществляют этот процесс совместно с клетками бобовых. Проникая в клетки корня растений и заражая их, бактерии вызывают образование клубеньков своеобразных опухолей. Клубеньковые бактерии в симбиозе с клетками корней бобовых могут фиксировать от 50 до 200 кг/га азота и больше, тогда как свободноживущие бактерии в почве фиксируют азота всего 1-6 кг/га. В данном случае и бобовые, и бактерии в принципе могут существовать один без другого, но при совместном взаимодействии эти виды достигают наибольшего процветания.
Мутуалистические взаимоотношения сложились также между водорослями и грибами в лишайниках. Водорослевым компонентом лишайника служит зеленая или сине–зеленая водоросль, а грибным – аскомицет (в состав тропических лишайников входят базидиомицеты). Фотосинтезирующая водоросль обеспечивает пищей оба компонента, а гриб защищает водоросль и снабжает её водой и минеральными солями. В мире известно около 10 000 видов лишайников. Грибы, входящие в лишайники, в отличие от водорослей обычно не могут жить самостоятельно. Поэтому гриб в известной степени выступает как паразит. Однако водорослям выгодна такая ассоциация, поскольку они получат возможность заселить многие места (поверхности голых камней, например, смытых склонов и т.д.), от освоения которых им пришлось бы отказаться без такого сожительства. Данный пример, как и ранее приведенные, является, очевидно, также доказательством параллельной эволюции организмов в сходных условиях и длительного их контактирования.
Краткий анализ ряда примеров мутуалистических отношений как наиболее важной части симбиоза раскрывает весьма сложную систему взаимодействий между различными организмами, обеспечивая им активность и процветание на земле в настоящее время.
Комменсализмом называют совместное существование различных организмов, при котором выгоду получает один вид, а для другого такое сожительство безразлично (нахлебничество). Примеры подобных отношений часто встречаются среди обитателей океана. Фактически, в каждой норе червя и в каждой раковине обитают незваные гости, находящиеся в организме хозяина, но не приносящие ему ни вреда, ни пользы. Наиболее ярким примером комменсализма является обитание маленькой рыбки в заднем кишечнике морского огурца (иглокожего). Рыбка сама проникает в хозяина и время от времени выходит наружу. Если бы она покинула приютившего хозяина, то стала бы жертвой других рыб.
Нейтрализм – отсутствие взаимодействий между видами.
Протокооперация – присутствие одного вида благоприятно для другого, но не является обязательным для его существования. Так, на панцирях многих крабов обитают различные кишечнополые, по–видимому, выполняющие роль маскировки. Это общество выгодно и кишечнополостному, который получает пищу, когда краб ловит и поедает других животных. Однако оба вида не находятся в абсолютной зависимости один от другого.
Антагонизм – отношения, при которых один или два вида испытывают ущерб (антибиоз, конкуренция, эксплуатация и т.д.).
Антибиоз – отношения между организмами, относящимися к разным видам, при которых один причиняет вред другому, не извлекая для себя из этих отношений видимого преимущества (не считая ослабления другого, вредного или конкурирующего вида). В качестве примера можно привести выделение вредных для других организмов веществ (капуста и виноград поэтому несовместимы).
Аменсализм – один вид угнетает другой, но не испытывает его влияния. Такое явление широко распространено между плесневыми грибами и бактериями. Многие плесневые грибы вырабатывают вещества, угнетающие развитие бактерий. Вероятно, это выгодно для гриба, устраняющего конкурента (бактерию) и получающего пищу в результате устранения последнего. Так, плесневый гриб Penicillum выделяет пенициллин, который подавляет рост бактерий. Применение бактерицидных агентов (антибиотиков) вызвало появление более частых заболеваний, обусловленных грибной инфекцией. В естественных условиях развитие болезнетворных бактерий сдерживается присутствием грибов. Уничтожение бактерий антибиотиками благоприятствует развитию патогенных грибов.
Паразитизм – один вид наносит ущерб другому и не может существовать без него, обитая на поверхности или внутри последнего. Классическим примером будет заразиха подсолнечника.
Хищничество – представители одного вида убивают и поедают представителей другого вида. Взаимоотношения между хозяином и паразитом, или хищником и жертвой не всегда вредны для хозяина или жертвы. Вредность таких взаимоотношений всегда ощущается в период их становления, а со временем их действие под влиянием сил естественного отбора ослабевает. Если губительное действие сохраняется, то паразит убивает хозяина и погибает сам, если не находит себе нового хозяина. Удивительный пример того, к чему приводит нарушение длительно существовавших взаимоотношений между хищником и жертвой, описан для Кайбабского плато на северном склоне Большого Каньона Колорадо. В 1907 г. в этом районе было около 4000 оленей и много хищников (пумы, волки). В результате отстрела хищников число оленей к 1925 г. увеличилось до 100000, что намного больше, чем могло прокормиться на имевшихся пастбищах. Олени поедали траву, сеянцы древесных пород и кустарников, нанося губительный урон растительности. В зимний период пищи не хватало, и в течение следующих зим большое число оленей погибло от голода. В итоге, популяция сократилась до 10000. Начальные взаимоотношения между хищниками и жертвой поддерживали устойчивое равновесие, при котором число оленей соответствовало наличным запасам на пастбище.
Анализ многих случаев отношений между хищником и жертвой, хозяином и паразитом показывает, что их взаимоотношения установились давно. В конечном итоге хищник или паразит не столь уж вредны, тогда как вновь появившиеся хищники или паразиты наносят большой вред. Особенно это характерно для сельскохозяйственных растений.
Для более глубокого понимания всех вопросов нужно хорошо знать образ жизни растении и животных, их роль в биоценозах, членами которых они являются. Растения и животные связаны более или менее тесным родством и, вместе с тем, находятся в различного рода межвидовых взаимоотношениях. Без обобщений подобного рода эволюционных связей и экологических взаимоотношений мир живых существ будет представлять для нас хаотическое скопление форм.
Конкуренция – организмы одного или разных видов действуют друг на друга в борьбе за пищу, местообитание и другие условия существования отрицательно. В качестве примера можно привести взаимоотношения, описанные Гаузе между популяциями простейшего Paramecium. При раздельном культивировании P. caudatum и P. aurelia и предоставлении им строго определенного количества пищи (бактерий) популяции размножались и достигали определенной численности. При совместном выращивании при том же ограниченном питании, к концу 16–го дня выживал только P. aurelia. Особи этого вида не нападали на представителей другого, не выделяли никаких веществ, они просто росли быстрее и поэтому брали верх в конкуренции за ограниченные запасы пищи (рис. 1).
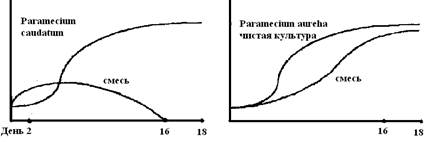
Рис. 1. Взаимоотношения между популяциями Paramecium
Конкуренция между организмами рассматривается на уровне внутривидовых и межвидовых взаимоотношений (все организмы конкурируют за многие ресурсы и трудно выделить определенные противоборствующие пары). Конкуренция проявляется всегда, когда экологические ниши организмов по каким–либо факторам пересекаются; она проявляется всегда за любые факторы, которых не достает для полного удовлетворения потребностей особей, популяций и видов в сообществе. Важнейшие факторы, за которые идет конкуренция, – это вода, свет, питательные вещества и кислород. В агросистемах на первых порах (после посева) между растениями конкуренция идет, в основном, за воду и минеральные вещества и лишь позже включается и за свет.
Иногда можно наблюдать одностороннее негативное воздействие одного вида по отношению к другому (явление аменсализма), хотя и не получающего от этой борьбы пользы самому себе (например, отношение между дерновинными злаками и амброзией на Кубани). Это явление – пример ассиметричной конкуренции, при которой влияние, например, амброзии можно не учитывать.
Между различными видами высших растений складываются сложные взаимоотношения, определяющие существование и состав популяций консортов. По В.Н. Сукачеву, следует выделить 3 основные группы взаимодействия между растениями: контактные, трансабиотические и трансбиотические.
Контактные делятся на две формы: а) со срастанием органов (симбиоз и паразитизм) и б) без срастания. Типичным примером первой формы является существование эпифитов на древесных растениях, срастание корней древесных растений, срастание стволов и др. Примером без срастания является воздействие друг на друга растущих рядом растений при их изгибах под действием силы ветра, воды и т.д.
Трансабиотические взаимоотношения являются определяющими между травянистыми растениями в ценозе. Их сущность заключается в том, что любое растение в процессе своей жизнедеятельности изменяет среду и тем самым оказывает влияние на соседствующее с ним растение. Это связано: 1) с использованием растением необходимых для него солнечной энергии, воды, углекислого газа, элементов минерального питания; 2) с выделением в окружающую среду продуктов его жизнедеятельности; 3) с отложением мертвого органического остатка (в почве и на поверхности); 4) с изменением водно-воздушного состояния почвы (задернением); 5) с созданием специфического фитоклимата. Среди всех выделенных факторов наибольшее значение имеет поглощение растениями воды и минеральных солей, перехват света, что вызывает их недостаток для других растений. В итоге возникает конкуренция за основные условия жизни. Например, для молодых посевов основное значение приобретает конкуренция за свет, возрастающая при увеличении сомкнутости и высоты стеблестоя. Она менее выражена на пастбищах при частом выпасе или скашивании и резче – на запущенных посевах сельскохозяйственных культур.
Совершенно не изучена конкуренция за кислород. Но вполне очевидно, что растения (особенно сорные), приспособленные к произрастанию в условиях недостатка кислорода, поглощают больше воды и питательных веществ в слабо аэрируемых почвах, чем не приспособленные к такой ситуации культурные виды.
Ч.Р. Дарвин борьбу за существование (конкуренцию) понимал в самом широком метафорическом смысле: борьба между особями одного вида (внутривидовая борьба) и между особями разных видов (межвидовая борьба), и с неблагоприятными условиями. Эти три типа борьбы за существование тесно связаны друг с другом.
Каждый вид своеобразен в конкуренции за факторы жизни, что определяется различиями в использовании ресурсов энергии и веществ отдельных горизонтов окружающей среды по периодам вегетации. Это связано с сезонным ритмом вегетации, величиной ассимиляционной и поглотительной поверхности и т.д. Отмеченные своеобразия усиливаются различиями консортивных связей. Например, бобовые, обеспечиваемые азотом клубеньковых бактерий, могут не выступать в качестве конкурентов с другими растениями за азот. Конкурентная способность зависит также от жизненного состояния особи, её потребности в отдельных элементах питания и т.д. Примером тому является выпадение из травостоев Западной Европы Bromus erectus и расширение присутствия Arrhenatherum elatius под влиянием внесения в почву в течение продолжительного времени больших количеств азотных удобрений.
Проведено много опытов для выяснения роли конкуренции за воду, элементы минерального питания (корневая конкуренция), свет с изоляцией корневых систем или надземных органов одних растений от других. Установлена большая роль корневой конкуренции во взаимоотношениях травянистых растений, что подтверждается опытами многих исследователей в различных районах мира. Установлено, что менее конкурентоспособные виды сильно подавляются соперничеством за азот более конкурентоспособных. Обычно конкуренция за азот проявляется жестче, чем за свет. Тем не менее, конкуренция за свет снижает площадь ассимиляции, уменьшает способность растений усваивать свет. Однако конкурентные взаимоотношения, как правило, сложные. Конкуренция за азот сопровождается конкуренцией за другие элементы питания, воду и т.д. При внесении высоких доз азота возрастает конкуренция между растениями за другие элементы питания.
Конкурентная способность вида – явление весьма сложное и определяется совокупностью морфологических и физиологических свойств растений (например, скоростью прорастания и скоростью роста, характером строения побегов и корневой системы, ритмом развития и продолжительностью жизни, способностью к возобновлению и размножению, количеством продуктивных семян и способом их распространения), а также требованиями к различным факторам их местообитания (температуре, свету, влажности, химическим факторам и т.д.). На сельскохозяйственных угодьях большую роль играет внутривидовая конкуренция. При весьма большом числе одновременно прорастающих растений отдельные их экземпляры сильно страдают от острой конкуренции, и их урожай резко падает. При широкорядном посеве конкуренция почти отсутствует, каждое растение развивается хорошо, но их малое число не обеспечивает больших урожаев. Возрастает также опасность зарастания сорняками. Для каждой культуры в определенных природно–климатических условиях эмпирически установлены нормы посева и посадки, позволяющие снимать достаточно хорошие урожаи. Внутривидовая конкуренция способствует сохранению вида, защищает его от конкуренции со стороны других видов (увеличение высоты и сомкнутости растений ограничивают рост и развитие других видов).
Межвидовая конкуренция проявляется в том, что малейшее изменение условий местообитания влечет за собой изменение количественных соотношений между ними и что одиночно растущие особи развиваются лучше всего на грядках, а в пределах растительного сообщества страдают от недостатка пищи. Это указывает на то, что растительное сообщество максимально использует все материнские ресурсы местообитания, не оставляя свободного пространства для видов, чуждых данному сообществу.
Различные виды растений характеризуются определенной экологией. Особенности их поведения можно установить только при совместном посеве. У растений следует различать физиологический и экологический оптимумы.
Конкурентные взаимоотношения культурных растений и сорняков. Формирование сорных растительных сообществ является отражением господствующих в данном месте абиотических условий существования и конкуренции, при которой одни виды вытесняют другие. Чтобы выдержать конкуренцию в отношении света, воды, минеральных веществ, жизненного пространства с окружающими растениями, у сорняков наряду с физиологической реакцией на внешние условия имеются также возможности, связанные с их конкуренцией, из которых наибольшее значение имеют продолжительность жизни, способ размножения, развитие корневой системы, способность к генерации, особенности роста и формирования вегетативных органов. Способность сорняков перерастать другие растения, расти под пологом, пронизывая корнями и корневищами другие растения, следует рассматривать как определенные формы борьбы за существование. Перерастание культурных растений быстро растущими сорняками – знакомая картина на полях. С другой стороны, рост многих сорняков подавляется быстро растущими и сильно затеняющими почву культурными растениями. Так, озимая рожь весьма эффективно подавляет проростки сорняков весной и осенью.
Большое значение имеет, какое растение из конкурирующих прорастает и будет развиваться первым. В зависимости от условий вегетации виды меняются, что хорошо просматривается при изучении конкуренции между сорняками и ячменем. Ячмень лучше поглощает азот почвы, чем ромашка. При избытке питания идет борьба за пространство между корневыми системами. В свою очередь, бухарник (Holcus mollis) поглощает азот лучше, чем ячмень. В этом случае преимущество остается за тем видом, который первый укоренился.
Мокрица (Stellaria media) ввиду более быстрого развития корней всегда имеет преимущество перед ячменем, поскольку в сообществе этих двух растений она быстрее использует азот. При любых условиях опытов всякому снижению продуктивности ячменя способствовало увеличение доли сорняков.
Трансбиотические взаимоотношения. Эта форма взаимодействия проявляется через влияние третьего организма, например, животных (млекопитающих, насекомых и т.д.), поедающих листья, плоды и другие части одних растений и не поедающих других. По причине ослабления через третий вид лучше поедаемые животными растения становятся менее конкурентоспособными. Особая роль в этой форме взаимоотношений принадлежит человеку (через внесение удобрений, поливы и другие его действия).
В опытах с душистым колоском и райграсом установлено, что на бедных почвах более конкурентоспособен душистый колосок, а на богатых – райграс. Однако на богатых почвах райграс сильно поражается и соотношение нередко меняется в пользу душистого колоска.
Подводя итог анализу взаимоотношений высших растений в агроценозах, следует заключить, что наибольшее значение в отношениях между ними имеет конкуренция за факторы жизни, в значительной степени определяемые особенностями их консортивных связей.
Остановимся подробнее на основных формах взаимоотношений. Внутривидовая конкуренция является важным фактором организации монодоминантных посевов. В абсолютно чистом виде этот вид конкуренции весьма ограничен во времени и в пространстве, поскольку трудно поддерживать посев без сорняков. Интенсивность внутривидовой конкуренции определяется плотностью посева, возрастом популяций, обеспеченностью организмов элементами питания, водой, режимом температуры, света и т.д.
При анализе внутривидовой конкуренции выделяется несколько общих закономерностей (Миркин, Злобин, 1990).
1. Конкуренция в посевах обостряется с возрастом растений: на ранних этапах формирования сообщества потребности организмов невелики и конкуренция вообще может не проявляться (наоборот, может проявляться эффект взаимопомощи). С усилением напряжения конкуренции выгоднее положение крайних (меньше контактирующих с конкурентом) особей.
2. Интенсивность конкуренции определяется пластичностью вида (у менее пластичных видов наблюдается изреживаемость, а у более пластичных при сохранении особей отмечается снижение роста и массы). В качестве примера можно привести посадки сосны и дуба в Ботаническом саду агроуниверситета (наблюдается изреживаемость древостоя на определенном возрастном отрезке за счет гибели части особей), а также участки самосева амброзии, где число особей в течение года остается постоянным, но их продуктивность сильно снижается.
3. Конкуренция регулируется плотностью особей и свойственна большинству живых организмов (увеличение массы растений коррелирует со снижением плотности популяции), что, безусловно, следует учитывать при выборе нормы посева, поголовья скота на пастбище и т.д. Например, на незасоренных полях норму высева можно снизить и получить высокий урожай за счет большой продуктивности хорошо развитых растений; на засоренных участках норма высева повышается, чтобы за счет загущения усилить давление на сорную растительность. На более поздних стадиях формирования посева наблюдается его самоизреживаемость и четко проявляется механизм зависимости плотности от смертности. При сочетании пластичности вида или сорта растения в затушенном посеве всегда будут менее развитыми, чем в разреженных. Самым консервативным признаком у большинства растений является масса семян; показатели кустистости, ветвления, числа цветков и т.д. сильно колеблются от условий. Самоизреживаемость растений в посевах в основном невысокая. Урожайность большинства культур выражается кривой, которая под действием человека может быть незначительно сдвинута вправо или влево, поэтому диапазон нормы их высева не очень широкий (рис. 2).

Рис 2. Влияние нормы высева на продуктивность посева.
Плотность посева и стеблестоя неодинаковы и непостоянны по годам и зависят от влажности, обеспеченности питательными веществами т.д. Низкая агротехника требует повышения нормы высева на 20–25%.
Вполне определенно просматривается разница между физиологической продуктивностью особи любой культуры (без конкуренции) и экологической (в посеве); последняя составляет 25–30% от максимальной. Чем выше агрофон, тем выше самоизреживание особей, свидетельствующее об усилении внутривидовой конкуренции, что экспериментально доказал еще В.Н. Сукачев (1928) в полевых опытах.
Внутривидовая конкуренция ускоряет (хотя и не всегда) развитие однолетников, и, наоборот, задерживает (тоже не всегда) развитие многолетников. Неодинаковость развития особей одной популяции объясняется тремя позициями: 1) неоднородностью популяции в генетическом отношении, 2) различиями условий развития особей популяции, 3) особи в силу каких–то причин (начиная с семени) различаются уровнем потенциальной жизнеспособности. Повышение внутрипопуляционного разнообразия (разное отношение к недостаткам различных условий, варьирование в конкурентоспособности, различия феноритмики, размеров и т.д.) позволяет популяции активнее расширять экологическую нишу и обеспечивает более эффективное использование его природных ресурсов. Повышать гетерогенность популяции можно посевами семян одного сорта в разные сроки, и незамоченными семенами, обработанными и необработанными микроэлементами и т.д. Иными словами, снижая конкуренцию, можно повышать урожаи, не вкладывая дополнительно энергоресурсы, а всего лишь оптимизируя агросистему.
Межвидовая конкуренция определяет характер формирования многовидовых посевов, например, отношение между культурными и сорными растениями. Этот тип конкуренции сложнее внутривидовой, поскольку различные организмы различаются конкурентоспособностью, потребностью к разным факторам местообитания, устойчивостью к вредителям, болезням, сорнякам и т.д. Изменениями условий местообитания в определенной степени можно менять отношения между видами (Работнов, 1984). Например, в условиях юга Таджикистана в посевах люцерны и голубого проса без удобрений особи люцерны в год накапливали I50 г сухой массы, а голубого проса всего лишь 18–20 г; при внесении азота под голубое просо масса люцерны снижалась относительно мало (до 100–112 г), но масса голубого проса повышалась до 140–160 г (для сравнения, в чистых посевах: люцерна без удобрений – 180 г, голубое просо с удобрениями – 210 г).
В полидоминантных сообществах конкурентоспособность молодых и старых особей весьма низкая, что и объясняет высокий процент выпадения в ювенильных и сенильных группах. Например, голубое просо на юге Таджикистана без удобрений держится в травостоях до 6 и даже 8 лет, а в травосмесях вытесняется через 3–4 года. На устойчивость растений в травосмесях влияет частота отчуждения: чем чаще срезается надземная масса растений, размножающихся семенами, тем быстрее вид уступает место другим, тем его конкурентоспособность снижается. Для вегетативно размножающихся видов это правило не подходит.
В культурных посевах контроль сорных растений осложняется их высокой конкурентоспособностью, более обильным запасом зачатков в почве, более широко экологической пластичностью. Для контроля сорных растений используют агротехнические приемы: своевременность выполнения всех технологических процессов, загущение посевов, расширение использования озимых и многолетних культур, существенно сдерживающих сорные растения. Необходимо знать сроки особой чувствительности отдельных культур к сорнякам: у зерновых – первый месяц вегетации, у свеклы – первые 2–4 недели вегетации и т.д.
Химические взаимоотношения включают воздействия выделений, накапливающихся в основном в почве (почвоутомление), на развитие различных организмов, а также влияние прямого выделения организмов на развитие других организмов.
Аллелопатическое почвоутомление обусловливается накоплением в почве биологически активных веществ до критического по токсичности уровня для особей собственного или другого вида, способствующее одностороннему развитию микрофлоры и усилению фитопатогенного фона. Аллелопатическое воздействие оказывают подвижные химические соединения, прежде всего фенольного типа (Henneguin, Juste, 1967). Химические воздействия растений способствуют их борьбе с сорняками. Например, ячмень меньше засоряется рядом сорняков, которые чувствительны к выделениям этой культуры – алкалоиду грамину (Оверленд, 1966). Весьма аллелопатически активны рожь, гречиха, конопля и др. Многолетнее выращивание люцерны способствует накоплению в почве сапонинов, сильно ингибирующих прорастание семян хлопчатника (Мишустин, Наумов, 1955; Степанова, 1975). Сильное почвоутомление вызывает люпин, бессменное выращивание которого приводит к его полному выпадению на третий – четвертый год (Гончарова и др., 1977). Вызывает осложнение и монокультура пшеницы, способствующая накоплению в почве фенолов (Muller–Wilmes, Loachke, 1980). Многие культуры (рис, кукуруза, табак, виноград, картофель и др.) мало утомляют почву и их можно возделывать по типу монокультуры. Наоборот, люцерна, сахарная свекла, горох, клевер являются мощными почвоутомителями.
Многие сорные растения отличаются аллелопатической активностью по отношению к культивируемым растениям. Аллелопатически агрессивны пырей ползучий (Грюммер, 1978), марь белая и росичка на кукурузу (Dayaday, Pons, 1978), а Dichanthium annulatum St. (корневые выделения, экстракты из стеблей) ингибируют рост проса африканского, капусты, салата (Dirvi, Hussain, 1979). Негативно влияют на многие растения щавель конский, выделяющий фенолы и влияющий на рост кукурузы и сорго, а из диких растений – на костер, пастушью сумку и т.д. Весьма активна в аллелопатическом отношении горчица салатная. Посевы лука в междурядьях картофеля и помидоров предохраняют их от заболевания фитофторой; капуста в междурядьях винограда угнетает его. Посев фасоли в междурядьях хлопчатника в Индии снижает инфекцию корневой гнили технической культуры грибами из рода Ризоктония. Посев клевера белого в чашки Петри с тимофеевкой и овсяницей дал меньше нормальных всходов, чем чистый, а у клевера лугового, наоборот (Ярошенко, Кушкова, 1971). В научной литературе приведены многочисленные данные о накоплении растениями веществ, влияющих на сорняки и другие растения, на всхожесть семян, рост растений, фиксацию азота микроорганизмами и т.д. (Раис, 1978). Подсев пижмы в междурядья картофеля в значительной мере снижает популяцию колорадского жука, который, как известно, очень устойчив к химическим веществам. Выделения гречихи и овса подавляют семенное размножение мари белой, а выделения полыни – размножение огурцов. Прорастание семян паразитного сорняка стриж наблюдается после получения ими биохимического сигнала о произрастании рядом культуры, к корням которой он сможет присосаться. При концентрации 1х10–16 стригон провоцирует прорастание и гибель до 50% семян стрижа. Иными словами, в искусственных сообществах, составленных видами различного происхождения, явление аллелопатии весьма нередкое и весьма впечатляющее.
Выделения растений оказывают существенное влияние на человека и животных. В случае пребывания в течение нескольких часов на поле цветущего кориандра, в зарослях багульника болотного и в непроветренной комнате с многочисленными цветами магнолии, ландыша, листьев эвкалипта человек испытывает головные боли и другие недомогания. Весьма сильное воздействие на организм человека оказывают летучие выделения сумаха ядовитого, лобелии одутлой, ясеница белого, которые вызывают потерю сознания, раздражение и даже ожоги кожи, сердцебиение и т.д. Другие растения (лимонник китайский, сосна и т.д.) выделяют летучие вещества, благоприятно действующие на человека.
Избирательно действие летучих выделений растений и на животных: выделения багульника болотного одуряюще действуют на собак; зеленики, чернокорень, кориандр, валериана, коровяк зопниковидный, черемуха и лавровишня выделяют вещества, изгоняющие мышей; пион обыкновенный, рута обыкновенная, бородавник отпугивают змей.
Весьма ощутимо влияние выделений растений на насекомых и других организмов. Так черемуха выделяет вещества, убивающие мух, комаров, мошку, слепней через воздух и через воду и отпугивающие вшей у животных. Подсев пижмы в междурядьях картофеля в значительной мере снижает популяцию колорадского жука.
Губительно влияют на тараканов полынь горькая, ромашка делегатская и персидская, живокость; на вошь платяную – семена пажитника, корни мерицы Лобеля и белой, цветы лаванды настоящей; на вошь головную – чеснок болотный; на тли – помидоры, настурция, паслен черный, ромашка, махорка, конопля; на клопа постельного – хрен, лавровишня, мухомор, клоповник, багульник болотный, молочай, чернокорень лекарственный, ромашка; на долгоносика амбарного – чеснок, бузина черная, конопля, кориандр, зеленик; на комаров – лавровишня, черемуха, эвкалипт; на блоху человеческую – лавровишня, полынь горькая, мята болотная, ромашка; на совку озимую – чеснок, черемуха; на муху домашнюю – рябина, ирга, черемуха, лавровишня, хрен, пижма обыкновенная; на моль шубную – махорка, лаванда настоящая, листья лимона, кипарисная трава; на плодоножку яблоневую – хрен; на белянку капустную – мята, розмарин; на муравьев – рябина, лавровишня, можжевельник, кипарисовик Лавсона, шалфей крупноцветковый; на клещей собачьих – лаванда, чабрец, шалфей, пиретрум; на паучного клещика – лук; на простейших – клюква обыкновенная.
Выделения растений по-разному влияют на другие растения. Озимая рожь угнетает озимую пшеницу; овес угнетает горох, люпин; вика и клевер угнетают овес; люпин угнетает картофель; нут угнетает картофель, помидоры, баклажаны, тыкву, огурец, дыню, арбуз, фасоль, подсолнечник, кукурузу, клещевину и кунжут; фасоль – пшеницу яровую; пшеница яровая – коноплю, горчицу, лен, анис; ячмень – люцерну синюю, нут, фасоль; горчица сизая – коноплю, нут; конопля – кенаф, нут; подсолнечник – клещевину, кукурузу; гречиха – кукурузу; помидоры – огурцы; лук – фасоль; репа – помидоры.
Положительные отношения складываются в смесях: нут – пшеница яровая, ячмень; фасоль – конопля, картофель, помидоры, баклажаны, суданка, чина, подсолнечник, тыква, дыня, арбуз, огурец, горох; горох – картофель, люцерна; картофель – ячмень; пшеница яровая – тыква, огурец, дыня, арбуз, соя; кукуруза – фасоль, нут.
В практической экологии весьма перспективно использование аллелопатической активности культурных растений к сорнякам, биологических методов защиты растений с применением природных выделений гормонов развития насекомых, половых аттрактантов и других веществ, управляющих поведением вредителей. Подсев крестоцветных и лекарственных растений к основной культуре способствует очищению поля от ряда сорняков, патогенов, вредителей, обогащает почвенную флору и фауну, а сами растения можно использовать на корм скоту или как фармсырье.
Смешанные посевы и посадки являются более продуктивными и устойчивыми, чем чистые посевы, но их широкое использование тормозится аллелопатической несовместимостью многих видов растений. Аллелопатические выделения растений обуславливают перенос химической информации, определяющей жизненную стратегию и тактику организмов и имеющую сигнальную природу.
Безусловно, в области химической экологии организмов необходимо продолжить исследования по физиологии и химизму растительных выделений в зависимости от экологических условий и взаимодействия с другими организмами, химизму превращений органического вещества в системе, аллелопатической активности и толерантности организмов, месту аллелопатии среди других форм взаимоотношений в устойчивости и продуктивности систем.
| Предыдущая |