
 Автор:
Автор: И.С. Белюченко, О.А. Мельник
Сельскохозяйственная экология
Учебное пособие. – Краснодар: Изд-во КГАУ, 2010. — 297 с.
| Предыдущая |
Содержание статьи:
4. Агроландшафт как экосистема
4.3. Консортивные связи
Помимо общеизвестных цепей питания целесообразно выделить в ценозах также сочетания разнообразных организмов, тесно связанных друг с другом в их жизнедеятельности известной общностью их судьбы (консортивная группа, или консорций). Такова, например, древесная порода со свойственными ей паразитами, сапрофитами, эпифитами (лишайники, мхи), симбионтами (микориза, микробы ризосферы и др.), вредителями, переносчиками пыльцы, семян и т.д. Без выделения и изучения консорциев наше знание биоценозов не будет полным и законченным (Раменский, 1952). В дальнейшем консорции стали рассматриваться как основные структурные части биоценоза и как элементарные энергетические системы, слагающие биоценозы.
Существует два подхода в выделении консорциев: 1) индивидуалистический, когда за основу консорциев берется особь, 2) популяционный, когда за основу принимается совокупность особей (популяция). Популяционный подход представляет большой интерес для агросистем, поскольку посевы отдельных сортов продовольственных или иных культур можно рассматривать как популяции.
Агроландшафты составлены большим числом различных видов организмов, имеющих между собой самые разнообразные взаимоотношения. Определенную роль в организации той или иной экосистемы имеют трофические связи, отражающие основное направление потока энергии в сообществе, использование одних организмов другими в качестве источника энергии. Отсюда возникло понятие о пищевых цепях. В качестве общей схемы пищевой цепи может быть такой ряд организмов: растение – фитофаг – зоофаг.
Взаимоотношения организмов в сообществах не ограничиваются только их трофическими связями. Например, паразиты и симбионты влияют на автотрофы не только тем, что заимствуют от них энергию, но и выделяют в их тела продукты своего метаболизма; гетеротрофы – фитофаги, контактно не связанные с автотрофами, влияют на последних, с одной стороны, используя их энергию и вещества и, с другой, меняя условия их существования (копытные не только поедают траву, но и уплотняют почву, оставляют несъеденными другие виды и т.д.). Велика для растений роль средообразующего влияния животных: животные не только используют плоды, пыльцу, нектар растений, но также способствуют опылению цветков растений, распространению их семян. Вместе с этим, именно животные нередко переносят бактериальные, вирусные и грибковые заболевания. В некоторых случаях, например, в случае поражения корней нематодами, возможно проникновение грибной инфекции в корни растений, стебли (боррер) и т.д.
Иногда трофические связи между организмами отсутствуют, а существуют только топические (связаны общим местом обитания). Например, древесные растения нередко используются эпифитами в основном как субстрат для прикрепления, а многими животными – как место для устройства гнезд. Иногда топические связи носят весьма своеобразный характер. Например, в Мексике и Ц. Америке наблюдается оригинальная связь между Acacia comigera и муравьем (Pseudomyrmex ferruginea). Муравьи образуют колонии во вздутых шипах акации и угнетают насекомых, поедающих акацию, тем самым защищая её от них. В отсутствие муравьев листья акации сильно объедаются другими насекомыми, что ведет к её отмиранию в течение 2–15 мес. Возникшие связи в результате сопряженной эволюции акации и муравьев привели к утрате акацией способности противостоять воздействию насекомых – фитофагов.
В связи с тем, что отношения между организмами в сообществах не замыкаются на трофике, представления о консорциях стали жизненно необходимыми. Если цепь питания показывает направление потока энергии и характер трансформации питательных веществ в биоценозах, то консорций отражает особенности взаимодействия между организмами в биоценозе.
В настоящее время консорции рассматриваются как основные структурные части естественных и искусственных биоценозов и как элементарные энергетические системы, слагающие биоценозы.
Структура консорция. В составе консорция выделяют центральный вид (детерминант) и консорты (совокупность организмов, зависимых от детерминанта энергетически или топически: паразитные растения, животные – эндобионты). В качестве детерминанта выступает ценопопуляция автотрофа. Основная часть консортов – гетеротрофы (исключая автотрофных эпифитов и полупаразитов). Консорты, связанные непосредственно с особями центрального вида, формируют первый концентр. Все они фитофага. Организмы, энергетически связанные с консорциями первого концентра, формируют второй и т.д.
Автотроф обеспечивает консорты первого концентра энергией и веществами и (или) предоставляет субстрат для их прикрепления или обитания, а также изменяет среду, в которой обитает. В понимании структуры консорция важнейшее значение имеют энергетические зависимости гетеротрофов от детерминанта. Гегеротрофы используют энергию и вещества, находящиеся в живых тканях автотрофов, в их отмерших органах, прижизненных выделениях. В соответствии с этим гегеротрофы первого концентра можно разделить на 3 группы организмов:
1) биогрофы, использующие энергию живых организмов;
2) сапротрофы, использующие энергию мертвых органов;
3) эккрисотрофы, использующие энергию прижизненных выделений.
Биотрофы потребляют органическое вещество, часто его минерализуют с образованием углекислого газа и воды, а у биотрофов – животных образуются также другие газы и соли. В результате работы сапротрофов образуются минеральные и органические вещества, некоторые из них очень стойкие, например, гумус почвы.
Таким образом, биотрофы и сапротрофы потребляют (консумируют) органические вещества и обеспечивают их частичную или полную минерализацию, т.е. являются редуцентами, минерализующими органические вещества. Различия между ними те, что одни используют живые, а другие – отмершие части растений.
Биотрофы, сапротрофы и эккрисотрофы представляют собою достаточно обособленные территориально и функционально группы организмов. Связанные с ними консорты второго и последующих концентратов также образуют относительно обособленные группы. Поэтому в консорциях можно выделить 3 направления энергетических связей: биотрофный, сапротрофный, эккрисотрофный.
Второй и последующие концентры состоят из биотрофов и сапротрофов. Биотрофы представлены животными – фитофагами, зоофагами и сапрофагами (грибы, актиномицеты, бактерии). Сапротрофы представлены в основном животными и грибами, использующими в качестве источника энергии тела отмерших животных, входящих в первый концетр консорция, а также части тела, периодически сбрасываемые при прохождении жизненного цикла, и экскременты более высокоорганизованных животных. Экскременты используются особой группой сапротрофов – копротрофами. Например, на пастбищах эта группа сапротрофов имеет большое значение.
Более высокоорганизованные животные также имеют своих консортов – эндо- и эпибионтов. Число концентров в биотрофной фазе невелико. В результате деятельности человека, уничтожившего в некоторых случаях хищных животных, число концентров ограничено нередко одним – двумя. Растительноядный слон завершает биотрофную фазу консорция, если не принимать во внимание его экто– и эндопаразитов, а вслед за насекомыми фитофагами может быть еще 2–3 и более концентров.
Животные – сапротрофы, питающиеся разлагающимися остатками растений в животных и экскрементами, потребляют вместе с органическим веществом большое количество организмов, участвующих в разложении органического вещества (грибы, бактерии), являющихся биотрофами. Тем не менее, сапротрофия имеет для них большее значение, чем биотрофия. Некоторые сапрофитные животные, например, дождевые черви, поедаются биотрофами (кротами), тем самым часть энергии и веществ включается из сапротрофной в биотрофную фазу. Это указывает на сложность энергетических связей в консорциях (рис. 3).
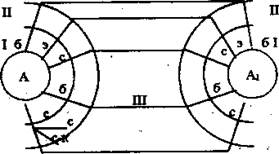
Рис. 3. Схема конвергенции энергетических связей.
А (автотроф), I, П – концентры; б – биотрофные связи, с – сапротрофные, э –эккрисотрофные, с–к – копротрофные (сапротрофные) связи.
Автотрофные растения в большей или меньшей степени своеобразны морфологически, анатомически, биохимически; своеобразны и их прижизненные выделения. Отмирающие органы автотрофов также различаются химически и механически, что определяет своеобразие в составе консортов первого концентра биотрофной, сапротрофной и эккрисотрофной структур консорция. Некоторые биотрофы входят в состав одной консорции близких автотрофов, у других биотрофов приуроченность к определенным консорциям не выражена, и они входят в состав многих консорций. При переходе из одного концентра в другой или с одного трофического уровня на другой происходит частичное рассеивание энергии (второй закон термодинамики) и наблюдается постоянная утрата энергетико-вещественной специфики консорции, а также постоянная конвергенция состава организмов определенных концентров в консорциях различных автотрофов. Кроме энергетических связей, являющихся вещественно–энергетическими (консорты получают от детерминанта вещества и энергию), между автотрофом и консортами существуют также трофические (вещественные) связи. Так, полупаразиты, например, погремок, способны к фотосинтезу и заимствуют от хозяина воду и минеральные вещества. Это растения – автотрофы энергетически, но гетеротрофы по отношению к воде и элементам питания.
Топически с детерминантами связаны только эпифиты (мхи, лишайники, водоросли), а также некоторые животные, устраивающие свои гнезда в стволах (дуплах), а также на ветвях деревьев. Возможно, что некоторые эпифиты частично используют в этой ситуации энергию и вещества детерминанта.
Основное ядро консорция образуется ценопопуляцией вида. В состав ценопопуляций входят особи различного возраста (от проростков до старых растений) и различного жизненного состояния (от угнетенных до хорошо развитых). Особи различного возраста и жизненного состояния отличаются по массе своих органов, химическому составу, морфологии, анатомии, что, безусловно, влияет на их использование биотрофами. Со старением увеличивается масса отмирающих органов и меняется химический состав. При отмирании состояния растений происходит повторное использование (реутилизация) содержащихся в них элементов питания растений (N, Р, К, Са и др.). Уровень их использования возрастает при низкой обеспеченности ими растений. Примерно то же самое происходит при старении или угнетении растений в связи с их неспособностью лучше использовать элементы минерального питания, имеющиеся в почве. Вариация химического состава отмерших органов растений оказывает прямое влияние на состав сапротрофов. Лишь с особями, имеющими генеративные органы, связана группа консортов, использующая плоды, семена, цветы. Численность группы зависит от доли генеративных особей в составе популяции. С возрастом растений изменяется приуроченность к ним грибной и другой паразитирующей микрофлоры. Различия габитуса надземной и подземной сферы детерминантов оказывают большое влияние на видовое разнообразие консортов. Это связано с тем, что в различных горизонтах среды формируются разные экологические ситуации.
Формирование консорции осуществлялось по мере становления автотрофов и сопряжено развивавшихся с ними организмов, способных с ними существовать. Становление консортов шло за счет местной флоры и фауны, способных существовать в определенных условиях экотопа. Только консортивно совместимые виды могут существовать вместе. Каждая из основных функциональных групп организмов, составляющих консорции (автотрофы, биотрофы, сапротрофы, эккрисотрофы), выполняет определенную функцию в ценозе. Консорции прямо и косвенно (через своих консортов) воздействует на среду обитания организмов в данном ценозе. Биотрофы оказывают влияние на продуктивность и соотношение автотрофов в фитоценозе, а также на обеспечение энергией и веществами сапротрофов и эккрисотрофов. Фитофаги сокращают поверхность усваивающих органов – листьев и корней автотрофов.
Фитопаразиты и связанные с автотрофами фитофаги угнетают детерминанты, выделяя в их ткани продукты своего метаболизма. Это снижает жизненное состояние автотрофов и интенсивность воздействия последних на среду. Фитофаги, конкретно не связанные с растениями, оказывают прямое влияние на среду: уплотняют, откладывают экскременты и т.д.
Грибы и бактерии широко представлены в экосистемах, и их симбиотрофная группа существенно влияет на конкуренцию автотрофов, обеспечивая их минеральными веществами. Особенно велика роль азотфиксаторов, активно влияющих на автотрофы и биоценоз в целом. Поступление азота ведёт к усилению минерализации. Отдельные симбиотрофы занимают промежуточное положение между биотрофами и сапротрофами. В частности, это относится к грибам, формирующим эктомикоризу, зависимым энергетически от органического вещества почвы и самого автотрофа. Эти консорты способны снабжать автотроф элементами минерального питания из органического вещества почвы без его минерализации.
Отдельные биотрофы опыляют цветки и распространяют плоды и семена, способствуя размножению видов (или их популяций). Большое значение имеют ризосферные микроорганизмы (зккрисотрофы). Они разнообразны по своим функциям. Велика их роль по детоксикации корневых выделений автотрофа, с которым они связаны, а также метаболитов сапротрофов. Бесспорно значение эккрисотрофов, влияющих на обеспечение азотом автотрофов. Сапротрофы минерализуют отмершие растения и животных, способствуют образованию доступных форм элементов минерального питания для автотрофов (особенно азота). В результате их жизнедеятельности образуются стойкие органические вещества (гумус), вещества, вступающие в химические реакции с минеральной частью почвы и воздействующие на автотрофы и их консорты, оказывая влияние на водно-воздушный режим (создание водопрочных структур, улучшение аэрации почвы, обогащение её органическим веществом и т.д.).
Автотрофы обеспечивают сапротрофы органическими веществами, что играет существенную роль в почвообразовании. Видовой состав консорции в нашем регионе изучен крайне слабо. Однако по имеющимся данным можно сделать заключение, что практически каждый растительный компонент агроландшафта имеет набор консортов, а отсюда – о большом количестве консортов.
Полночленность консории. Автотроф и консорты могут иметь экологически различные ареалы. В основных ареалах автотроф имеет, скорее всего, постоянные и дополнительные консорты. Попадание вида в новое сообщество и освоение им новой ситуации сопровождается формированием его консорций. В состав консорция нового автотрофа входят виды, свойственные этому ценозу, а также те, которые перекочевали сюда вместе с автотрофом. Если местная флора и фауна не имеют видов, свойственных для внедряющегося вида, то образуется немногочисленные консорции, возникающие при интродукции автотрофа за пределы его ареала. В качестве примера можно назвать опунции, завезенные в Австралию из Америки в конце прошлого столетия для создания живых изгородей. Условия вегетации оказались благоприятными, а в фауне не нашлось видов, способных стать для опунции эффективными биотрофными консортами. Поэтому она быстро распространилась на обширных площадях и обусловила снижение продуктивности пастбищ. Аналогичный пример с интродукцией бобового кустарника марабу на Кубу из США. При разработке мер борьбы с ним оказалось более целесообразным устранить неполночленность их консорций путем завоза насекомых – эффективных биотрофных консортов.
По-другому сложилась судьба с интродукцией подсолнечника в Россию. Он начал поражаться заразихой, которая до того паразитировала на местных формах полыни. На родине (Северная Америка) в числе консортов подсолнечника заразихи не было. В новом ареале в составе консорций автотрофа появился биотроф, шатающий его урожай. Со временем селекционеры вывели заразихоустойчивые сорта.
Нередко консорции отдельных автотрофов полночленны (в их составе находятся все местные организмы). Однако считать эти консорции полностью полночленными нельзя, поскольку в других районах могут быть организмы, способные входить в их состав. Занос таких видов, способных стать эффективными биотрофными консортами местных растений, может сопровождаться катастрофическими последствиями, поскольку эволюция автотрофа протекала не сопряжено с заносным биотрофом, и он не обладает соответствующими защитными реакциями, обеспечив возможность совместного успешного обитания. Например, в северо-восточных штатах США местный вид каштана (Castanea dentate) во второй половине прошлого века широко встречался в лесах и, наряду с дубом, был преобладающим. В 1904 г. было впервые отмечено поражение каштана занесенным из Китая паразитным грибом Endothia parotitis. В восточных областях Китая этот вид паразитирует на местных видах каштана, но не наносит им существенного вреда, что указывает на сопряженность их эволюции. В США гриб быстро распространился, что сопровождалось гибелью каштана. В течение 20 лет каштан погиб в Новой Англии, а в последующие 20 лет поражение каштана грибом распространилось до южных границ его произрастания в штате Джорджия (Costing, 1956). Дубово-каштановые и дубово-каштаново-гикориевые леса стали дубовыми и дубово-гикориевыми.
Все эти примеры указывают на большое практическое значение полночленности консорция. Иногда человек относительно легко решает эту проблему. Уничтожение барбариса (промежуточный хозяин линейной ржавчины – Puccinia granjinis) привело к неполночленности консорций пшеницы и способствовало поднятию её урожайности. Человек выключает нежелательные консорты и вводит новые, которые способствуют повышению продуктивности автотрофов, например, применение нитрагина (бактериальное удобрение) способствует усилению роста бобовых и поднятию их урожайности.
Таким образом, изучение консорциев как составных структур экосистем имеет большое хозяйственное и, безусловно, теоретическое значение.
| Предыдущая |