
 Автор:
Автор: А.С. Замотайлов, И.Б. Попов, А.И. Белый
Экология насекомых
Краткий курс лекций. – Краснодар: КубГАУ, 2009. – 184 с.
| Предыдущая |
Содержание статьи:
Тема 2. Введение в популяционную экологию
2.3. Пространственная структура популяций
По Тимофееву-Ресовскому под популяцией понимается совокупность особей определенного вида, в течение достаточно длительного времени (большого числа поколений) населяющих определенное пространство, внутри которого практически осуществляется та или иная степень панмиксии и нет заметных изоляционных барьеров, которая отделена от соседних таких же совокупностей особей данного вида той или иной степенью давления тех или иных форм изоляции. Экологический аспект популяционной биологии направлен на изучение жизни популяции как формы существования вида в составе конкретных экосистем.
Пространственная структура популяций выражается характером размещения особей и их группировок по отношению к определенным элементам ландшафта и друг к другу и отражает свойственный виду тип использования территории. Пространственная структурированность определяет наиболее эффективное использование ресурсов среды (пищевых, защитных, микроклиматических и др.), снижая хаотичность их использования и, как следствие, уровень конкурентных взаимоотношений между особями внутри популяции. В свою очередь, пространственная структура определяется особенностями пространства и биологии данного вида.
Различают три типа пространственного распределения особей в популяции: равномерный (регулярный), диффузный (случайный) и агрегированный (групповой, мозаичный) (рис. 4).
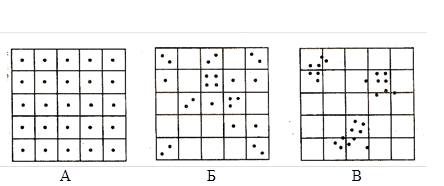
Рисунок 4 – Типы пространственного распределения особей в популяциях (по Шилову, 1997):
А – равномерное; Б – диффузное; В — мозаичное
Равномерный тип распределения в идеале характеризуется равным удалением каждой особи от всех соседних; величина расстояния между особями соответствует порогу, за которым начинается взаимное угнетение. Таким образом, теоретически этот тип распределения в наибольшей степени соответствует задаче полного использования ресурсов при минимальной степени конкуренции, однако в действительности этот тип распределения в природе встречается достаточно редко. Близким к этому характер распределения встречается в уплотненных популяциях некоторых тлей, располагающихся на одновидовых зарослях растений или в плотных кулигах саранчовых. Практическая невозможность реализации этого идеального типа распределения определяется, прежде всего, неоднородностью среды обитания.
Диффузный тип распределения особей встречается в природе значительно чаще, при нем особи распределены в пространстве неравномерно, случайно. Такой тип распределения характерен для подавляющего количества видов насекомых, что определяется неравномерным распределением ресурсов по поверхности биотопа. Именно этот тип характерен и для фитофагов, и для сапрофагов, и для хищников с паразитами.
Агрегированный (мозаичный) тип распределения выражается в образовании группировок особей, между которыми остаются достаточно большие незаселенные территории. Биологически это связано либо с резкой неоднородностью среды, либо с выраженной социальной структурой, действующей на основе активного сближения особей. Это особенно характерно для социальных видов насекомых, например, муравьев, термитов, ос, шмелей и медоносной пчелы. В случае неравномерности среды подобное распределение характерно, например, для насекомых моноксилофагов, питающихся каким-либо одним видом мертвой древесины.
Для насекомых как подвижных в большинстве своем животных ведущее значение в определении характера пространственной структуры популяции имеет степень привязанности к территории. В крайних вариантах, встречающихся у насекомых достаточно редко, это свойство выражается либо оседлым, либо кочевым образом жизни, для большинства видов характерны промежуточные варианты. В то же время благодаря тому, что преимагинальные стадии насекомых – личинки и куколки – как правило, менее подвижны, чем взрослые и сильнее привязаны к источнику пищи или месту зимовки, то они занимают меньшую территорию, хотя и сильнее от нее зависимы. Взрослые же стадии могут перемещаться (мигрировать) за пределы областей, внутри которых возможно развитие полного цикла вида.
Оседлый образ жизни в его крайнем варианте характерен для многих видов общественных насекомых (муравьев, термитов), семьи которых в течение многих поколений и, соответственно, иногда десятков лет занимают определенную территорию, площадь и конфигурация которой определяются особенностями пространства и давления конкурентов того же вида. В силу всего этого она может несколько изменяться, но, в целом, представляет собой территорию обитания одной семьи, куда представители конкурирующих семей того же вида не допускаются. Маркировка территории осуществляется за счет выделения дюфуровых желез, расположенных на голове, протоки которых открываются возле челюстей. Секрет этих желез наряду с выделениями ядовитой железы, расположенной на конце брюшка рабочих особей и солдат муравьев достаточно длительное время способен сохраняться на субстрате, к которому прикасаются маркирующие территорию особи.
Пример крайнего варианта кочевого образа жизни можно также встретить у муравьев – обитателей тропических областей, которых так и называют – муравьи-кочевники. Эти общественные насекомые не имеют определенной территории, не строят гнезд, в которых выращивается потомство, а постоянно перемещаются в неопределенном направлении колонной, в центре которой находится матка-царица, рабочие особи с расплодом, перетаскиваемым постоянно в челюстях. По периметру такой колонны располагаются муравьи-солдаты, охраняющие матку и рабочих особей с расплодом, а также занимающиеся разведкой и охотой. Подобные колонны муравьев находятся в постоянном движении и останавливаются только при достижении каких-либо препятствий, например, реки или для кормления.
Большинство других насекомых занимают промежуточное положение между этими двумя крайними вариантами. Например, даже для территориальных насекомых имеют место ситуации, как правило, связанные с размножением, когда часть колонии мигрирует на определенное расстояние с целью поиска места для основания новой колонии, например, роение семей медоносных пчел.
Очень распространены миграции насекомых, связанные с поиском источников пищи, когда плотность популяции увеличивается слишком сильно и родная стация не способна ее прокормить. Особенно это характерно для ряда видов саранчовых, которые способны формировать многомиллиардные стаи взрослых особей, которые перелетают на большие расстояния в поисках пищи, или кулиги личинок последних возрастов. Элементы подобного поведения встречаются у многих видов тлей, формирующих крылатых самок-расселительниц, основная функция которых состоит в формировании новых колоний на соседних растениях.
Широко известны сезонные миграции бабочки монарха Danaus plexipus из Северной Америки, которая совершает ежегодные миграции с восточного побережья США и Канады на зимовку в Калифорнию и острова Мексиканского залива. Причем эти миграции каждый раз совершает новое поколение бабочек и совершает единственный раз. В Краснодарском крае встречается олеандровый бражник Daphnis nerii, обитающий на Черноморском побережье в районе Сочи. Его гусеницы выкармливаются на листьях произрастающего здесь олеандра, однако, зимующие куколки как правило не переживают даже небольших отрицательных температур, и на следующий год местная популяция бражника возникает за счет мигрантов из Турции. Имаго олеандрового бражника встречали даже в Финляндии, за 2,5 тысячи километров от мест произрастания кормовых растений. Большинство же видов насекомых могут перемещаться лишь на очень небольшие расстояния, и их цикл развития связан, как правило, с очень небольшим участком территории, зачастую единственным растением.
| Предыдущая |